Voltage-Gated Ion Channels
Cells have to import and export ions in order to survive. Since ions are charged molecules, they do not pass through the cell membrane. This means they have to be transported by proteins that are embedded in the membrane.
There are many different kinds of transporters. Some of them use up energy to move ions across the membrane by a process called active transport. Some of them simply open a channel and allow ions to pass through. Let's look at some channel proteins.
A typical channel protein consists of several subunits arranged around a central hole that permits passage of ions. In this case we'll be dealing with ion channel proteins that allow movement of Na+ (sodium) or K+ (potassium) ions from the outside of the cell (top) to the cytoplasm (bottom). The part of the protein that spans the membrane is usually an α helical region. In fact, there are usually several α helices in a channel protein.
The flow of ions must be regulated, so these channel proteins must be capable of opening and closing to allow or restrict the flow of Na+ or K+. This is why they're called gated channels. There are several ways of controlling the gate. We are going to be discussing mechanisms that respond to the membrane potential―the difference in charge on either side of the membrane. For example, the wave of membrane depolarization that occurs as a nerve impulse, or action potential, moves along the axon of a nerve cell.
These channel proteins are called voltage-gated ion channels.
The simplest kind of voltage-gated ion channels are the K+ (potassium) channels found in bacteria and in eukaryotes. The role of potassium channels is to allow influx of potassium if the concentration inside the cell drops below some minimal value. Normally the concentration of K+ in the cytoplasm is about 100 times greater than the concentration outside the cell because K+ ions are actively transported into the cell as Na+ ions are pumped out. The concentration of Na+ outside the cell is about 10 times higher than the inside concentration.
From time to time, these concentrations are perturbed (e.g., during the firing of a nerve impulse) and the sodium and potassium channels will open to restore the resting state concentrations. The gates in the ion channels will only open for 1 millisecond but this is enough to allow entry of thousands of ions per channel.
The structures of several bacterial K+ channels are known and recently the structure of the homologous channel from rat was solved. The structures are very similar. Let's look at the rat voltage-gated K+ channel (Longe et al. 2005a; Long et al. 2005b).

The complete channel consists of four identical subunits colored red, green, yellow, and blue in the figure on the left. The crystallized protein has an additional β subunit at the bottom attached to the T1 cytoplasmic domain but I've removed it in order to simplify the description. The β subunit contains an oxidoreductase activity that's coupled to the influx of potassium.
If you look at the structure of the single red subunit you can see that it consists of a bundle of four transmembrane α helices labelled S1-S4 on the left and two others (S5 and S6) on the top right. The S5 and S6 helices are the ones that form the central channel when the four subunits come together. This is most easily seen in the top view at the bottom of the figure. The black dot in the middle represent a K+ ion about to pass through the channel.
The structure shown is the open conformation that allows passage of ions. The closed conformation was modeled from the known structure of the bacterial channels and experimental data that implicated the S4-S5 linker helix in the gating process. You can see this linker helix in the figure above. It joins the S1-S4 bundle to the S5, S6 bundle. It is not one of the transmembrane helices. The results are shown below.
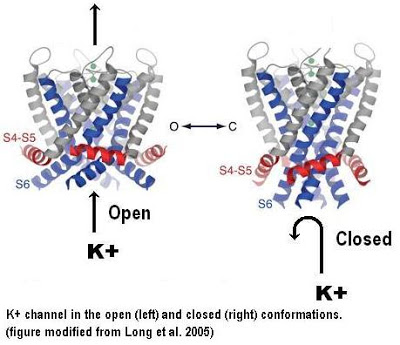
In the closed conformation, the S4-S5 helix responds to changes in the membrane potential by twisting from a horizontal position to a slightly more vertical position. This causes the S5 and S6 transmembrane helices to also adopt a more vertical orientation. Since the same conformational change occurs in all four subunits, the net effect is to make the entire protein more narrow and this closes the channel in the middle of the protein. The switch between the open and closed conformations is extremely rapid and it is very sensitive to changes in the membrane potential.
Long, Stephen B., Campbell, Ernest B., MacKinnon, Roderick (2005a) Crystal Structure of a Mammalian Voltage-Dependent Shaker Family K+ Channel. Science 309:897-903.
Long, Stephen B., Campbell, Ernest B., MacKinnon, Roderick (2005b) Voltage Sensor of Kv1.2: Structural Basis of Electromechanical Coupling. Science 309:903-908.

